Although Mutants That Do Not Produce Exotoxins Are Relatively Easy to Isolate
To search the entire book, enter a term or phrase in the form below Custom Search Endotoxins are part of the outer membrane of the cell wall of Gram-negative bacteria. Endotoxin is invariably associated with Gram-negative bacteria whether the organisms are pathogenic or not. Although the term "endotoxin" is occasionally used to refer to any cell-associated bacterial toxin, in bacteriology it is properly reserved to refer to the lipopolysaccharide complex associated with the outer membrane of Gram-negative pathogens such as Escherichia coli, Salmonella, Shigella, Pseudomonas, Neisseria, Haemophilus influenzae, Bordetella pertussis and Vibrio cholerae. The relationship of endotoxin (lipopolysaccharide) to the bacterial cell surface is illustrated in Figure 1 below. The biological activity of endotoxin is associated with the lipopolysaccharide (LPS). Toxicity is associated with the lipid component (Lipid A) and immunogenicity is associated with the polysaccharide components. The cell wall antigens (O antigens) of Gram-negative bacteria are components of LPS. LPS elicits a variety of inflammatory responses in an animal and it activates complement by the alternative (properdin) pathway, so it may be a part of the pathology of Gram-negative bacterial infections. In vivo, Gram-negative bacteria probably release minute amounts of endotoxin while growing. This may be important in the stimulation of natural immunity. It is known that small amounts of endotoxin may be released in a soluble form by young cultures grown in the laboratory. But for the most part, endotoxins remain associated with the cell wall until disintegration of the organisms. In vivo, this results from autolysis, external lysis mediated by complement and lysozyme, and phagocytic digestion of bacterial cells. Compared to the classic exotoxins of bacteria, endotoxins are less potent and less specific in their action, since they do not act enzymatically. Endotoxins are heat stable (boiling for 30 minutes does not destabilize endotoxin), but certain powerful oxidizing agents such as superoxide, peroxide and hypochlorite, have been reported to neutralize them. Endotoxins, although antigenic, cannot be converted to toxoids. A comparison of the properties of bacterial endotoxins and classic exotoxins is shown in Table 1. Table 1. Characteristics of bacterial endotoxins and classic exotoxin 
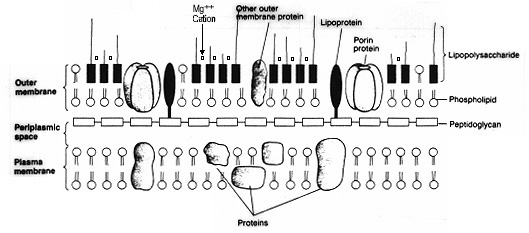
Figure 1. Structure of the cell envelope of a Gram-negative bacterium (e.g. E. coli). The peptidoglycan sheet is actually in the periplasm, the space between the inner (plasma) and outer membranes. The inner face of the outer membrane is composed of phospholipid, the same as in the plasma membrane. The outer face of the outer membrane contains some phospholipid but primarily it is composed of lipopolysaccharide which has the amphipathic qualities of a phospholipid. Lipopolysaccharide is endotoxin. Imbedded in the outer membrane are various outer membrane proteins, lipoprotein and porin proteins, which sometimes play a role in virulence of a Gram-negative bacterium, but they are not considered endotoxins.
PROPERTY ENDOTOXIN EXOTOXIN CHEMICAL NATURE Lipopolysaccharide (mw = 10kDa) Protein (mw = 50-1000kDa) RELATIONSHIP TO CELL Part of outer membrane Extracellular, diffusible DENATURED BY BOILING No Usually ANTIGENIC Yes Yes FORM TOXOID No Yes POTENCY Relatively low (>100ug) Relatively high (1 ug) SPECIFICITY Low degree High degree ENZYMATIC ACTIVITY No Often
PYROGENICITY Yes Occasionally
The Role of LPS in the Outer Membrane of Gram-negative Bacteria
The function of the outer membrane of Gram-negative bacteria is to act as a protective permeability barrier. The outer membrane is impermeable to large molecules and hydrophobic compounds from the environment. LPS is essential to the function of the outer membrane. First, it establishes a permeability barrier that is permeable only to low molecular weight, hydrophilic molecules. In the E. coli the ompF and ompC porins exclude passage of all hydrophobic molecules and any hydrophilic molecules greater than a molecular weight of about 700 daltons. This prevents penetration of the bacteria by bile salts and other toxic molecules from the GI tract. It also a barrier to lysozyme and many antimicrobial agents. Second, in an animal host, it may impede destruction of the bacterial cells by serum components and phagocytic cells. Third, LPS may play a role as an adhesin used in colonization of the host. Lastly, variations in LPS structure provide for the existence of different antigenic strains of a pathogen that may be able to bypass a previous immunological response to a related strain.
Chemical Nature of Endotoxin
Most of the work on the chemical structure of endotoxin has been done with species of Salmonella and E. coli. LPS can be extracted from whole cells by treatment with 45% phenol at 90o. Mild hydrolysis of LPS yields Lipid A plus polysaccharide.
Lipopolysaccharides are complex amphiphilic molecules with a mw of about 10kDa, that vary widely in chemical composition both between and among bacterial species The general architecture of LPS is shown in Figure 2. The general structure of Salmonella LPS is shown in Figure 3 and the complete structure of Salmonella lipid A is illustrated in Figure 4.

Figure 2. General architecture of Lipopolysaccharide
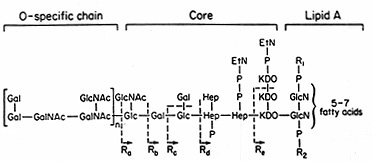
Figure 3. General Structure of Salmonella LPS. Glc = glucose; GlcNac = N-acetyl- glucosamine; Gal = galactose; Hep = heptose; P = phosphate; Etn = ethanolamine; R1 and R2 = phoshoethanolamine or aminoarabinose. Ra to Re indicate incomplete forms of LPS. The Rd2 phenotype (not shown) would have only a single heptose unit. The Rc, Rd2, and Rd1 mutants lack the phosphate group attached to Hep.

Figure 4. Complete structure of the Lipid A Moiety of LPS of S. typhimurium, S. minnesota, and E. coli
LPS consists of three components or regions: Lipid A, an R polysaccharide and an O polysaccharide.
Region I. Lipid A is the lipid component of LPS. It contains the hydrophobic, membrane-anchoring region of LPS. Lipid A consists of a phosphorylated N-acetylglucosamine (NAG) dimer with 6 or 7 fatty acids (FA) attached. Usually 6 FA are found. All FA in Lipid A are saturated. Some FA are attached directly to the NAG dimer and others are esterified to the 3-hydroxy fatty acids that are characteristically present. The structure of Lipid A is highly conserved among Gram-negative bacteria. Among Enterobacteriaceae Lipid A is virtually constant.
The primary structure of Lipid A has been elucidated and Lipid A has been chemically synthesized. Its biological activity appears to depend on a peculiar conformation that is determined by the glucosamine disaccharide, the PO4 groups, the acyl chains, and also the KDO-containing inner core.
Region II. Core (R) antigen or R polysaccharide is attached to the 6 position of one NAG. The R antigen consists of a short chain of sugars. For example: KDO - Hep - Hep - Glu - Gal - Glu - GluNAc -
Two unusual sugars, heptose and 2-keto-3-deoxyoctonoic acid (KDO), are usually present, in the core polysaccharide. KDO is unique and invariably present in LPS and so it has been used as an indicator in assays for LPS (endotoxin).
With minor variations, the core polysaccharide is common to all members of a bacterial genus (e.g. Salmonella), but it is structurally distinct in other genera of Gram-negative bacteria. Salmonella, Shigella and Escherichia have similar but not identical cores.
Region III. Somatic (O) antigen or O polysaccharide is attached to the core polysaccharide. It consists of repeating oligosaccharide subunits made up of 3 - 5 sugars. The individual chains vary in length ranging up to 40 repeat units. The O polysaccharide is much longer than the core polysaccharide, and it maintains the hydrophilic domain of the LPS molecule. A major antigenic determinant (antibody-combining site) of the Gram-negative cell wall resides in the O polysaccharide.
Great variation occurs in the composition of the sugars in the O side chain between species and even strains of Gram-negative bacteria. At least 20 different sugars are known to occur and many of these sugars are characteristically unique dideoxyhexoses, which occur in nature only in Gram-negative cell walls. Variations in sugar content of the O polysaccharide contribute to the wide variety of antigenic types of Salmonella and E. coli and presumably other strains of Gram-negative species. Particular sugars in the structure, especially the terminal ones, confer immunological specificity of the O antigen, in addition to "smoothness" (colony morphology) of the strain. Loss of the O specific region by mutation results in the strain becoming a "rough" (colony morphology) or R strain.
The elucidation of the structure of LPS (Figure 3) relied heavily on the availability of mutants each blocked at a particular step in LPS synthesis. The biosynthesis of LPS is strictly sequential. The core sugars are added sequentially to Lipid A by successive additions, and the O side chain is added last, one preassembled subunit at a time. The properties of mutants producing incomplete LPS molecules suggests the nature and biological functions performed by various parts of the LPS molecule.
In E. coli and Salmonella, loss of the O antigen results in partial loss of virulence, suggesting that this portion of LPS is important during a host-parasite interaction. It is known that such "rough" mutants are more susceptible to phagocytosis and serum bactericidal reactions.
Loss of the more proximal parts of the core, as in "deep rough" mutants (i.e. in Rd1, Rd2, and Re mutants in Figure 3) makes the strains sensitive to a range of hydrophobic compounds, including antibiotics, detergents, bile salts and mutagens. This area contains a large number of charged groups and is thought to be important in maintaining the permeability properties of the outer membrane.
Mutants in the assembly of Lipid A cannot be isolated except as conditional lethal mutants and this region must therefore be essential for cell viability. The innermost region of LPS, consisting of Lipid A and three residues of KDO, appears to be essential for viability, presumably for assembling the outer membrane.
LPS and virulence of Gram-negative Bacteria
Both Lipid A (the toxic component of LPS) and the polysaccharide side chains (the nontoxic but immunogenic portion of LPS) act as determinants of virulence in Gram-negative bacteria.
The O polysaccharide and virulence
Virulence, and the property of "smoothness", is associated with an intact O polysaccharide, The involvement of the polysaccharide chain in virulence is shown by the fact that small changes in the sugar sequences in the side chains of LPS result in major changes in virulence. How are the polysaccharide side chains involved in the expression of virulence? There are a number of possibilities:
1. O-specific antigens could allow organisms to adhere specifically to certain tissues, especially epithelial tissues.
2. Smooth antigens probably allow resistance to phagocytes, since rough mutants are more readily engulfed and destroyed by phagocytes.
3. The hydrophilic O polysaccharides could act as water-solubilizing carriers for toxic Lipid A. It is known that the exact structure of the polysaccharide can greatly influence water binding capacity at the cell surface.
4. The O antigens could provide protection from damaging reactions with antibody and complement. Rough strains of Gram-negative bacteria derived from virulent strains are generally non virulent. Smooth strains have polysaccharide "whiskers" which bear O antigens projecting from the cell surface. The O antigens are the key targets for the action of host antibody and complement, but when the reaction takes place at the tips of the polysaccharide chains, a significant distance external to the general bacterial cell surface, complement fails to have its normal lytic effect. Such bacteria are virulent because of this resistance to immune forces of the host. If the projecting polysaccharide chains are shortened or removed, antibody reacts with antigens on the general bacterial surface, or very close to it, and complement can lyse the bacteria. This contributes to the loss of virulence in "rough" colonial strains.
5. The O-polysaccharide or O antigen is the basis of antigenic variation among many important Gram-negative pathogens including E. coli, Salmonella and Vibrio cholerae. Antigenic variation guarantees the existence of multiple serotypes of the bacterium, so that it is afforded multiple opportunities to infect its host if it can bypass the immune response against a different serotype. Furthermore, even though the O polysaccharides are strong antigens, they seldom elicit immune responses which give full protection to the host against secondary challenge with specific endotoxin.
Lipid A and virulence
The physiological activities of LPS are mediated mainly by the Lipid A component of LPS. Lipid A is a powerful biological response modifier that can stimulate the mammalian immune system. During infectious disease caused by Gram-negative bacteria, endotoxins released from, or part of, multiplying cells have similar effects on animals and significantly contribute to the symptoms and pathology of the disease encountered.Since Lipid A is embedded in the outer membrane of bacterial cells, it probably only exerts its toxic effects when released from multiplying cells in a soluble form, or when the bacteria are lysed as a result of autolysis, complement and the membrane attack complex (MAC), ingestion and killing by phagocytes, or killing with certain types of antibiotics.
The injection of living or killed Gram-negative cells or purified LPS into experimental animals causes a wide spectrum of nonspecific pathophysiological reactions, such as fever, changes in white blood cell counts, disseminated intravascular coagulation, hypotension, shock and death. Injection of fairly small doses of endotoxin results in death in most mammals. The sequence of events follows a regular pattern: (1) latent period; (2) physiological distress (diarrhea, prostration, shock); (3) death. How soon death occurs varies on the dose of the endotoxin, route of administration, and species of animal. Animals vary in their susceptibility to endotoxin.
The mechanism is complex. In humans, LPS binds to a lipid binding protein (LBP) in the serum which transfers it to CD14 on the cell membrane, which in turn transfers it to another non-anchored protein, MD2, which associates with Toll-like receptor-4 (TLR4). This triggers the signaling cascade for macrophage/endothelial cells to secrete pro-inflammatory cytokines and nitric oxide that lead to characteristic "endotoxic shock". CD14 and TLR4 are present on several cells of the immunological system cells, including macrophages and dendritic cells. In monocytes and macrophages, three types of events are triggered during their interaction with LPS:
1. Production of cytokines, including IL-1, IL-6, IL-8, tumor necrosis factor (TNF) and platelet-activating factor. These, in turn, stimulate production of prostaglandins and leukotrienes. These are powerful mediators of inflammation and septic shock that accompanies endotoxin toxemia. LPS activates macrophages to enhanced phagocytosis and cytotoxicity. Macrophages are stimulated to produce and release lysosomal enzymes, IL-1 ("endogenous pyrogen"), and tumor necrosis factor (TNFalpha), as well as other cytokines and mediators.
2. Activation of the complement cascade. C3a and C5a cause histamine release (leading to vasodilation) and affect neutrophil chemotaxis and accumulation. The result is inflammation.
3. Activation of the coagulation cascade. Initial activation of Hageman factor (blood-clotting Factor XII) can activate several humoral systems resulting in
a. coagulation: a blood clotting cascade that leads to coagulation, thrombosis, acute disseminated intravascular coagulation, which depletes platelets and various clotting factors resulting in internal bleeding.
b. activation of the complement alternative pathway (as above, which leads to inflammation)
c. plasmin activation which leads to fibrinolysis and hemorrhaging.
d. kinin activation releases bradykinins and other vasoactive peptides which causes hypotension.
The net effect is to induce inflammation, intravascular coagulation, hemorrhage and shock.
Textbook of Bacteriology Index
etheridgediespithe.blogspot.com
Source: https://textbookofbacteriology.net/endotoxin.html
0 Response to "Although Mutants That Do Not Produce Exotoxins Are Relatively Easy to Isolate"
Publicar un comentario